| Zurück zur Übersicht | (Veröffentlicht mit
freundlicher Genehmigung von Dr.
W-E Lönnig) |
Wolf-Ekkehard Lönnig:
Last Update 5. August 2010: neu ist der Teil 2 ab p. 113 (frühere Updates zum ersten Teil siehe p. 112).Einige zumeist kleinere orthographische Korrekturen am 13. 12. 2011.
"Die Affäre Max Planck", die es nie gegeben hat
Diffamierungspolitik, weltanschauliche Motivation und (Berufsverbots-)Ziel der AG Evolutionsbiologie
Teil 2
Einige Ergänzungen und Literaturangaben zu beiden Teilen
| Zurück |
Teil II / 3 von 4 ab
Seite 123 |
zum Kapitel
12 |
(4) UK: "Charles Darwin (1875) hat als einer der ersten Naturforscher die Entwicklung der Fangblasen von Utricularia analysiert und gezeichnet. Seine Ergebnisse, die in Zusammenarbeit mit Sohn Francis erarbeitet wurden, sind in Abb. 10.2 zusammengefasst."
Abbildung aus Kutschera 2007, p. 289:
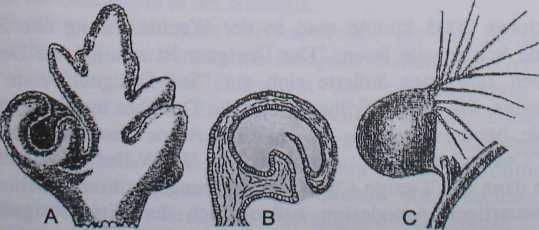
"Abb. 10.2: Entwicklung der Saugfalle beim Wasserschlauch (Utricularia vulgaris). Junges Blatt aus einer Winterknospe, mit einer entstehenden Fangblase (A); junge Saugfalle im Querschnitt (B) und funktionstüchtiges Organ mit reizbaren Tasthaaren (C) (Nach Darwin 1875)." (Kutscheras Abb. 10.2 ist aus 3 getrennten Abbildungen aus Darwins Werk von 1875 zusammengesetzt; dort C zuerst.)
(4) W-EL: Turionen (Winterknospen) haben nach Troll und Dietz (1954) keine Schläuche bzw. keine bereits "entstehenden Fangblasen" (siehe ihren Kommentar in der ausführlichen Fußnote zum Punkt (5) sowie Maier 1973 und Adamec 1999, 2010), sodass sich Darwins Abbildung (A) möglicherweise auf das Blatt eines jungen Triebes aus einer Winterknospe bezieht oder was wahrscheinlicher ist auf einer Fehlinterpretation beruht (Darwin selbst räumte diese Möglichkeit ein).
Kutschera lässt weiter unerwähnt (wie übrigens auch MN 2009), dass Darwin gar nicht erkannt hatte, dass es sich um einen aktiven Fangvorgang handelt. Dazu erinnere ich wie folgt an den hier geringfügig veränderten Kommentar aus http://www.weloennig.de/Utricularia2010.pdf p. 15/16:
Darwin schreibt (1875, Insectivorous Plants pp. 405, 406, 409, vgl. http://darwin
online.org.uk/content/frameset?viewtype=text&itemID=F1217&pageseq=1):
"Animals enter the bladders by bending inwards the posterior free edge of the valve, which from being highly elastic shuts again instantly. " As I felt much difficulty in understanding how such minute and weak animals, as are often captured, could force their way into the bladders, I tried many experiments to ascertain how this was effected. Worauf die ausführliche Beschreibung seiner vielen Versuche dazu folgt. Ergebnis: "To ascertain whether the valves were endowed with irritability, the surfaces of several were scratched with a needle or brushed with a fine camel-hair brush, so as to imitate the crawling movement of small crustaceans, but the valve did not open. Some bladders, before being brushed, were left for a time in water at temperatures between 80° and 130° F. (26°.6-54°.4 Cent.), as, judging from a widespread analogy, this would have rendered them more sensitive to irritation, or would by itself have excited movement; but no effect was produced. We may, therefore, conclude that animals enter merely by forcing their way through the slit-like orifice; their heads serving as a wedge. It is difficult to conjecture what can attract so many creatures, animal- and vegetable-feeding crustaceans, worms, tardigrades, and various larvae, to enter the bladders. Mrs. Treat says that the larvae just referred to are vegetable-feeders, and seem to have a special liking for the long bristles round the valve, but this taste will not account for the entrance of animal-feeding crustaceans. Perhaps small aquatic animals habitually try to enter every small crevice, like that between the valve and collar, in search of food or protection.
Die soweit bekannt ersten Beschreibungen Utricularias als karnivore Pflanze stammen von Lobel (1591) und Holland (1868). James and Patricia Pietropaolo (2001, p. 123) kommentieren die Historie der Entdeckung des Fangmechanismus von Utricularia wie folgt:
"The elucidation of the carnivorous plant habit of this genus began to unfold when Cohn, in 1857, discovered that they captured Perch fry. Both Cohn and Darwin thought that the prey pushed the trap door open, entered, and when the door was closed found themselves entrapped. It was Mary Treat, who in 1876, discovered that the prey did not swim into the trap, but rather were sucked in when the trap was set off and thereby captured.
Der Fangvorgang ist also wesentlich komplexer als sich das Darwin und Wallace (nach Einsicht in Darwins Werk) seinerzeit vorgestellt hatten (siehe den Vorschlag von Wallace zum damaligen Kenntnisstand der Fallenfunktion von Utricularia in den nächsten Absätzen). Entsprechend steigt auch der Schwierigkeitsgrad einer adäquaten Erklärung (vgl. http://www.weloennig.de/Utricularia.html und vor allem zur hochkomplexen Anatomie der Struktur der Verschlussklappe von Utricularia sowie deren Funktion gemäß den Studien Guttenbergs unter http://www.weloennig.de/Utricularia_H_von_Guttenberg.pdf ).
Wallace hob in seinem Brief an Darwin vom 21. Juli 1875 (siehe http://darwin online.org.uk/ ) einige Schwierigkeiten zur Entstehung der Saugfalle von Utricularia durch die natürliche Selektion hervor (obwohl er ihm zunächst beruhigend versicherte: " I daresay there is no difficulty") und bat Darwin, diese Punkte doch einmal direkt anzusprechen (nun hatte Darwin wie oben dokumentiert noch nicht das Prinzip der aktiven Saugfalle erkannt: " animals enter merely by forcing their way through the slit-like orifice; their heads serving as a wedge " was Wallace erst gemäß unserem heutigen Wissenstand über Utricularia sagen würde, bleibt natürlich offen):
"Dear Darwin,Many thanks for your kindness in sending me a copy of your new book [Insectivorous Plants]. Being very busy I have only had time to dip into it yet. The account of Utricularia is most marvellous, and quite new to me. I'm rather surprised that you do not make any remarks on the origin of these extraordinary contrivances for capturing insects. Did you think they were too obvious? I daresay there is no difficulty, but I feel sure they will be seized on as inexplicable by Natural Selection, and your silence on the point will be held to show that you consider them so! The contrivance in Utricularia and Dionæa, and in fact in Drosera too, seems fully as great and complex as in Orchids, but there is not the same motive force. Fertilisation and cross-fertilisation are important ends enough to lead to any modification [das ist zwar auch nicht zutreffend], but can we suppose mere nourishment to be so important, seeing that it is so easily and almost universally obtained by extrusion of roots and leaves? Here are plants which lose their roots and leaves to acquire the same results by infinitely complex modes! What a wonderful and long-continued series of variations must have led up to the perfect "trap" in Utricularia while at any stage of the process the same end might have been gained by a little more development of roots and leaves, as in 9,999 plants out of 10,000! Is this an imaginary difficulty, or do you mean to deal with it in future editions of the "Origin"? Believe yours very faithfully, Alfred R. Wallace.
Mir ist nicht bekannt, dass Darwin darauf geantwortet hat, d. h. er hat die Herausforderung ("I feel sure they will be seized on as inexplicable by Natural Selection, and your silence on the point will be held to show that you consider them so!) nicht angenommen (auch nicht in "future editions of the "Origin"", denn er konnte diese Fragen so wenig befriedigend beantworten, wie das der Darwinismus heute nach mehr als 135 Jahren der weiteren Forschung kann. Und die Schwierigkeiten haben zugenommen.
- (5) UK: "Im Herbst bilden die Wasserschlauch-Sprosse am Ende große Knospen, die abfallen und den kalten Winter überdauern. Die jungen Blätter, welche diese Knospen umschließen, enthalten Vorstufen von Saugfallen (Abb. 10.2 A, B)."
- (5) W-EL: Wir müssen hier zwei Dinge deutlich unterscheiden, nämlich die Entwicklung aus Turionen (die "großen Knospen, die abfallen", den Winter überdauern und die sich nach Troll und Dietz allgemein durch Schlauchlosigkeit auszeichnen 2 1) und die normale Ontogenese, d. h. die Entwicklung der Pflanzen aus Samen.

Links: Turio von Utricularia vulgaris aggr. (Foto Kristian Peters). Rechts: Austreibende Turio von U. bremii (Foto Denis Barthel (vgl. http://de.wikipedia.org/wiki/Datei:Utricularia_vulgaris_agg_turion.jpeg und http://de.wikipedia.org/wiki/Wasserschl%C3%A4uche , last update Juni 2010)
{kind=link}
Nach allen mir bislang bekannten Daten zeigen erst die sich entwickelnden Blätter an den Trieben der Turionen die Bildung von funktionsfähigen Saugfallen (siehe die Abbildung oben rechts), nicht aber die Turionen selbst.
- (6) UK: "Während der Ontogenese der Utricularia-Blätter wandeln sich diese Anlagen [sic!] in funktionstüchtige Fangblasen um (Abb. 10.2 C), die somit als Blatt-Metamorphosen (-Abwandlungen) zu interpretieren sind (Darwin 1875)."
- (6) W-EL: Die reguläre Ontogenese aus Samen verläuft z. B. wie folgt:
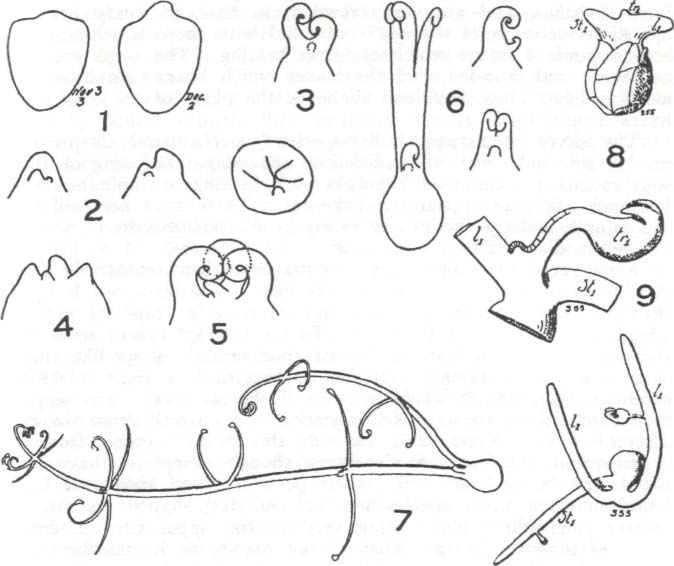
Aus Lloyd 1942/2007, p. 221: "Fig. 6. 1-7, Utricularia purpurea. 1, Early stages of germination of a seed (3.3 mm long) from which only one growing point arose (to he followed by others later in all probability); 2, Three figures in a row, three views of an early stage of germination, a later stage of which is shown in 3, in which two young growing points show circination; 4 and 5, Two following stages in the germination of a seed which produced three growing points all of nearly the same age, with a fourth, secondary to the middle growing point; 6, A case in which fasciation occurred, the two figures on the right show an early stage of germination, a much later stage is shown on the left, in which it is seen that one of the growing points had divided, an abnormality; 7, An advanced stage in germination (15 mm long), one of the three growing points still quiescent; five whorls of branches (the maximum seen) were produced, as shown on the longer stolon of this figure. At the first whorl of this, the bud of a branch stolon of indefinite growth is seen. No traps are produced on the two dorsal branches of the third and fourth whorls.
8-10, Ulricularia cleistogama. 8, Early stage of germination showing primary stolon and primary leaf, with the primary trap on the leaf near its base; g, A later stage in detail showing the origin of the trap from the leaf base; 10, A more advanced stage of the seedling in which a second leaf arose in the place of a primary trap."
Kommen wir jetzt (wieder) zur Ontogenese der Saugfallen an sich: Nach Haeckels "Biogenetischem Grundgesetz" würde man erwarten, dass die Ontogenese die Phylogenese widerspiegelt (adulte Stadien sollten in der Individualentwicklung durch Addition immer weiterer Stadien bis zum heutigen Phänotyp vorkommen). Nehmen wir Haeckel beim Wort, so ist Folgendes festzustellen: Keine der vielen ontogenetischen Entwicklungsstufen hat in diesem mehr oder weniger kontinuierlichen Bildungsprozess bis zur Vollendung der komplexen Saugfalle irgendeine Funktion (auch hier gilt: "Erst mit dem Endeffekt wird der Nutzen für den Daseinskampf erreicht, nicht aber mit irgendeiner Entwicklungsstufe"). Damit entfallen für sämtliche ontogenetischen Bildungsstufen auch irgendwelche Selektionsvorteile. Unterbricht man die Ontogenese des Fangapparates an irgendeiner der hundert und mehr möglichen Stellen vor Vollendung der Entwicklung, erhält man immer funktionslose Stadien (oder schärfer formuliert: funktionswidrige Zwischenstufen), d. h. einen unvollendeten Apparat, der nicht in der Lage ist "über das Einsaugen und Verdauen kleiner Tiere, die weitgehend aus stickstoffreichen Proteinen aufgebaut sind ... [den Bedarf des Wasserschlauchs] am Mangelelement N" zu decken (soweit dieser Stickstoffbedarf der entscheidende Punkt ist).
Interpretiert man die ontogenetischen Entwicklungsstufen als Evolutionsstadien der Vorzeit, so müsste man schlussfolgern, dass der gesamte hypothetische jahrmillionenlange Evolutionsprozesses über "unzählige aufeinander folgende kleine Mikroevolutionsschritte" nichts mit Selektion zu tun hatte, sondern völlig funktionslos und autonom verlaufen wäre. Die bei mir 2010, p. 14 erwähnten Schlauchrudimente hingegen kosten Energie und tragen, soweit mir bekannt ist, nichts zur Ernährung des Wasserschlauchs bei.
Überdies kommen die von mancher Seite behaupteten Zwischenstufen, die speziell die Fallen von Pinguicula und Genlisea auszeichnen oder zumindest diesen in charakteristischen Merkmalen stark geglichen und z. T. auch so funktioniert haben sollen, in der Ontogenese Utricularias überhaupt nicht vor, abgesehen von der generellen Interpretation der Genlisea-Falle als epiascidiates Blatt. Diese Interpretation wird jedoch auch sonst bei allen karnivoren pitchers (krug-, kannen-und schlauchförmigen Fallen) vorgenommen. Da dieser Punkt von einigen Autoren übermäßig im Sinne einer evolutionären Deutung betont wird, erhebt sich die Frage: Wie sonst sollte die Ontogenese in allen diesen Fällen beginnen?
Pinguicula ist nach Auffassung der meisten mir bekannten Autoren die "primitivste" Gattung der Lentibulariaceen 2 2 und von sehr ähnlichen Formen will man evolutionär die anderen beiden Gattungen, Genlisea und Utricularia, herleiten. Nach der "Biogenetischen Grundgesetz" (heute meist nur noch als "Regel" bezeichnet) könnte man ein mehr oder weniger flächige Anfangs- (oder geringfügig spätere) Stadien in der Entwicklung der Fallen der beiden abgeleiteten Gattungen erwarten. Das ist jedoch nicht der Fall.
Ein Pinguicula-ähnliches Stadium fehlt völlig in der Ontogenese (Individualentwicklung) der Fangapparate von Genlisea und Utricularia.
Es wäre auch ziemlich widersinnig, erst ein Klebfallenstadium (oder auch nur eine rudimentäre Klebfalle) in der Entwicklung der Fangapparate dieser Gattungen aufzubauen, um dann dieses (bzw. diese) in die völlig anderen (ihnen typisch eigenen) Fangvorrichtungen umzubauen. Aber, so wendet vielleicht mancher Leser ein, gibt es da nicht doch starke Ähnlichkeiten in der Ontogenese von Genlisea und Utricularia? Sehen wir uns also ein paar wesentliche Daten aus der Entwicklung der Fangapparate dieser beiden Gattungen zur Verdeutlichung des oben Gesagten etwas näher an.
Juniper, Robins and Joel schreiben 1989, pp. 66/67 zum Start der Individualentwickung des Utricularia-Fangapparates :
"The opening of the trap arises in the very young plants as a slit caused by the invagination of the rounded primordium."
Bei Genlisea hingegen sehen bereits die ersten Entwicklungsstadien deutlich anders aus (Juniper et al. p. 72):
"Marginal growth leads to the formation of an invagination in the tip of the cylindrical primordium of the trap. In contrast to the spherical invagination in the primordial trap of Utricularia (see page 66), the invagination in Genlisea is tubular, arising from an extended marginal growth.
Auffällig ist zunächst, dass nicht einmal eines der postulierten vielen Übergangsstadien vom normalen zum ascidiaten Blatt in der Individualentwicklung wiederholt wird. Die Ontogenese der Fangapparate beginnt statt dessen direkt wie oben zitiert mit "a slit caused by the invagination of the rounded primordium" (Utricularia), bzw. "the formation of an invagination in the tip of the cylindrical primordium of the trap" (Genlisea).
Die Ontogenese Utricularias verläuft weiter wie folgt (Juniper et al. p. 67):
"The lips of the slit turn inwards, the upper becoming the trap door while the lower lip becomes the threshold (Meierhofer, 1902). The sides of the entrance then extend, moving the lips apart to produce a funnel-shaped vestibule around the opening."
Folgendes ist dagegen bei Genlisea zu beobachten (p. 72) (wobei die evolutionären Homologiedeutungen der Autoren denkbar fragwürdig sind und selbst unter der Voraussetzung des Darwinschen Weltbilds ebenso gut als Konvergenzen interpretiert werden könnten):
"While the basal portion of the invagination develops into the sub-spherical hollow utricle, its neck forms a long tube which gradually widens towards the mouth [long tube is totally missing in Utricularia] where it forms a transverse slit with two lips (see Fig. 4.17C), a shorter ventral inner lip (homologous to the trap door in Utricularia) and a dorsal lip which is arched over the opening (homologous to the threshold in Utricularia traps).
Die evolutionären Homologiedeutungen 2 3 der Lippen setzen einen gemeinsamen Vorfahren nicht nur mit einem Mund voraus, sondern auch mit einer annähernd "shorter vertral inner lip", einer Lippe, die im Sinne der Abstammungslehre mit der Tür der Utricularia-Falle homolog sein soll, und womöglich auch mit "a dorsal lip which is arched over the opening", die als homolog mit dem Widerlager (threshold) der Saugfalle gedeutet wird. Eine genauere Untersuchung dieser Frage scheint mir nicht unangebracht zu sein. Wie auch immer, von solchen gemeinsamen Vorfahren ist nichts bekannt.
Überdies sieht auch der Rest der von Juniper et al. beschriebenen Entwicklung des Fangapparates bei Genlisea völlig anders aus als bei Utricularia. Die Autoren schreiben weiter auf p. 72:
"In the last phase of the trap development [in Genlisea], cell divisions are restricted to the two sides of the mouth only, where the lips are connected. These cell divisions lead to the elongation of the two parallel lips. Two arms thus develop on top of the tube, circinate when young and fully expanded when mature. The two parallel lips form spirals along the arms. The helical arms, with their spirals turning in opposite directions (Fig. 4.17) are interconnected on top of the tube. In this manner the slit, which extends in both arms, forms an extremely long and narrow mouth through which small animals can enter the trap (Fig. 4.17A, B).
Da der extrem lange und schmale Mund in regelmäßigen Abständen durch schmale Seitenstege unterbrochen wird (vgl. die Abbildung in Lönnig 2010, p. 42), handelt es sich in Wirklichkeit um zahlreiche kleinere Münder.
Für Utricularia hingegen weisen die Autoren dagegen auf folgende Punkte der auch wieder völlig unterschiedlichen weiteren Ontogenese hin (p. 67) 2 4:
"Although contiguous at their lateral extremities, the door and the threshold [which are not formed in Genlisea at all] differ in their anatomical characters (Figs. 4.125 and 4.14). The threshold becomes semicircular and forms a massive thickening, which preserves the shape of the opening and resists any crumpling when the trap is set. The side walls of the trap are thinner where they articulate with the threshold so that no distortion is exerted on the surrounding tissue when the side walls bend as the trap sets (Fig. 4.10,5). In transverse section, the threshold is roughly triangular and continuous with the lower trap wall. The upper, free surface of the threshold shows at least three distinct regions: the inner zone, made of epidermal cells, forming a shelf projecting into the interior of the trap; the outer zone, doorstep, continuous with the lateral walls of the entrance; and between them is the middle zone, which is the 'pavement epithelium' on which the door lies when the trap is closed. The pavement epithelium fits precisely the shape of the lower door edge and is provided with special devices which prevent leakage of water into the trap when the trap is set."
Angesichts dieser grundlegenden, systematischen Unterschiede in der Ontogenese (Individualentwicklung) der beiden Gattungen mit entsprechend so unterschiedlichen Endergebnissen (vgl. Tabelle, Lönnig 2010, p. 43) erscheint mir der einleitende Satz der Autoren zur Entwicklung der Genlisea-Falle ("The development of the Genlisea trap closely resembles that of Utricularia: both are epiascidiate leaves") nur auf dem Hintergrund ihres evolutionären Weltbilds verständlich.
Mit dem Argument der epiascidiate leaves könnte man ebenso Folgendes behaupten: "The development of the Genlisea trap closely resembles those of Heliamphora, Sarracenia, Darlingtonia, Nepenthes and Cephalotus: all these are epiascidiate leaves." Da die letzteren auch nach der Evolutionstheorie unabhängig von Genlisea und die Sarraceniaceen wiederum unabhängig von den Nepenthaceen und beide unabhängig von Cephalotaceen entstanden sind, warum sollte man dann die unabhängige Entstehung der so erstaunlich unterschiedlichen Fangapparate von Genlisea und Utricularia von vornherein ausschließen und stattdessen einen Abstammungszusammenhang suggerieren?
Zu makroevolutiven Ansätzen vgl. meinen Beitrag Goethe, Sex and Flower Genes (1994), Schwartz 2005, sowie Lönnig 2010, pp. 109-116.
| Zurück |
Ende Teil II / 3 von 4 |
zum Kapitel
12 |
| Zurück zur Übersicht |